Foundations
What is the Sigma-1 Receptor?
A strange protein inside cells, why DMT binds to it, and the increasingly serious question of what that binding might mean
1. What this article is
This is the closing piece in the Foundations series. The previous five — DMT, Ayahuasca, 5-MeO-DMT, Harmine and the MAOIs, and What is the Default Mode Network? — built up the substance pharmacology and the cognitive-neuroscience vocabulary that this site spends most of its time engaged with. This one introduces a target that gets mentioned in passing throughout that earlier material but has not been properly explained: the sigma-1 receptor.
The piece is more technical than the previous five. It deals with intracellular biology rather than substance experience, with chaperone proteins and stress responses rather than ceremony or molecular comparison. The reward for working through it is access to a strand of psychedelic-pharmacology research that the surface-receptor story (5-HT2A, 5-HT1A, the receptors covered in the earlier primers) does not capture, but that may turn out to be doing more than the surface story can account for on its own.
A short author note, in the same third-person register as the DMN essay. The writer's engagement with the sigma-1 receptor is as a reader of the literature rather than as a participant in any practice that involves it directly — though the long-term cellular effects of DMT and 5-MeO-DMT use, which the writer does have direct experience of, may well be mediated in part through this receptor. The science is open enough that this is plausible rather than established.
2. The discovery
The sigma-1 receptor has one of the stranger origin stories in contemporary pharmacology. It was discovered not because anyone was looking for it, but because a class of drugs bound to something that was not a known receptor and someone eventually had to name the something.
In the 1970s, researchers studying opioid pharmacology noticed that certain compounds — N-allylnormetazocine and related benzomorphans — produced effects in animals that resembled some opioid effects but differed from others. The compounds bound to a protein in the brain that initially appeared to be an unusual opioid receptor subtype. The protein was provisionally named the "sigma receptor" (σ-receptor) and was, for several years, classified as a member of the opioid receptor family.
This turned out to be wrong on every count. By the late 1980s, careful pharmacology had established that the sigma receptor was not an opioid receptor at all — opioid antagonists like naloxone did not block its effects, and its tissue distribution and binding profile were inconsistent with the opioid family. The name persisted by inertia. The protein was reclassified as a "non-opioid sigma site" and remained pharmacologically mysterious for another decade.
Further confusion followed. Researchers identified what appeared to be two distinct sigma receptors — sigma-1 and sigma-2 — based on differences in their binding profiles and tissue distribution. Sigma-1 was eventually purified, sequenced, and cloned in 1996, and its structure determined in detail over the following years. Sigma-2 turned out to be a different protein entirely, finally identified molecularly only in 2017 (it is now known to be the TMEM97 protein), confirming that the two sigma subtypes were unrelated despite sharing a name.
So the sigma-1 receptor, as currently understood, is the molecular entity originally identified through 1970s opioid research, identified as non-opioid in the 1980s, structurally characterised from the 1990s onward, and now understood to be a distinct kind of protein with no significant relationship to either opioid receptors or to "sigma-2." The naming history is a mess. The biology has turned out to be considerably more interesting than the naming history suggested.
3. What kind of protein it is
To understand why the sigma-1 receptor matters, it helps to start with where it lives.
Most receptors that drugs are designed to interact with are cell-surface receptors — proteins embedded in the outer membrane of cells, with one part facing the outside world and another part facing the cell's interior. A molecule arriving at the cell surface binds to the external-facing part of the receptor, and this binding triggers signals that propagate inward through the cell. Almost every major class of psychiatric and neurological drug — antidepressants, antipsychotics, opioids, classical psychedelics — works by binding to cell-surface receptors of one kind or another.
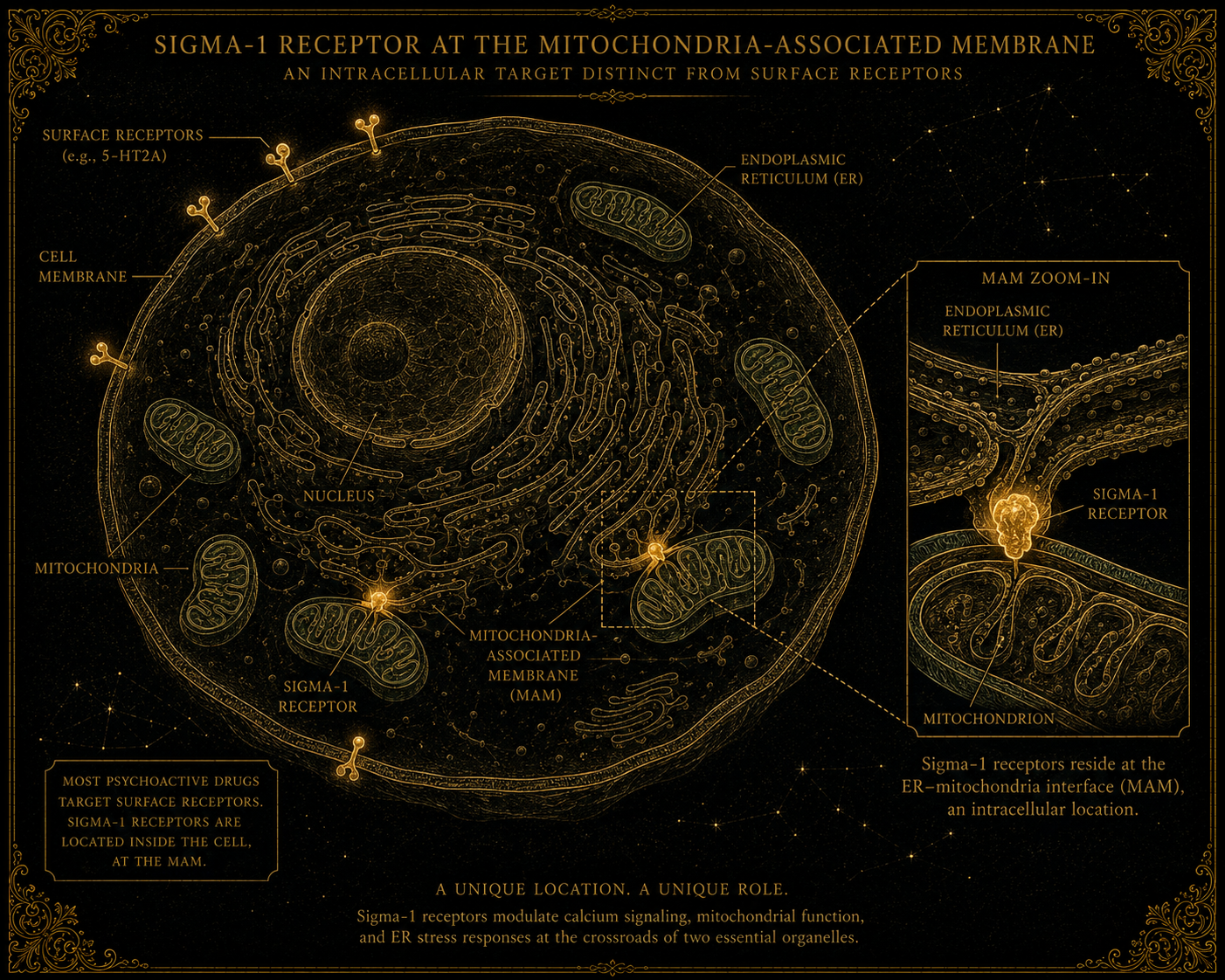
The sigma-1 receptor is different. It is not on the cell surface. It lives inside the cell, embedded in the membrane of an internal organelle called the endoplasmic reticulum (ER). More specifically, it sits at the junction between the endoplasmic reticulum and the mitochondria — a contact zone called the mitochondria-associated membrane (MAM), where these two organelles meet and exchange information.
This location is unusual. Most receptors live in places that respond to signals from outside the cell. The sigma-1 receptor lives in a place that responds to signals from inside the cell — to cellular stress, to imbalances in calcium signalling, to demand on the protein-folding machinery, to disturbances in the mitochondrial energy supply.
The protein itself is small (about 223 amino acids), has a relatively simple structure (it crosses the membrane just once), and behaves more like a chaperone than like a conventional receptor. A chaperone protein, in cellular biology, is a protein whose job is to assist other proteins — helping them fold correctly, stabilising them under stress, sometimes shepherding them to their correct locations within the cell. The sigma-1 receptor seems to operate primarily in this chaperone-like role, modulating the behaviour of other proteins it associates with rather than driving signalling cascades of its own.
This combination — intracellular location, chaperone-like function, sensitivity to cellular stress — makes the sigma-1 receptor a different kind of pharmacological target from the surface receptors that dominate medicine. Drugs that bind to it are doing something at a different layer of the cell, with consequences that propagate through cellular processes rather than through neural signalling alone.
4. What it does
The functional role of the sigma-1 receptor has emerged in stages over the last twenty years, and is still being worked out. The following is the current consensus picture, with the appropriate caveats that some of this remains contested.
Calcium signalling regulation. The endoplasmic reticulum stores calcium ions and releases them in regulated bursts that drive much of cellular signalling. The sigma-1 receptor associates with proteins that control calcium release from the ER and modulates their behaviour. When cells are under stress, sigma-1 activity changes in ways that protect against calcium-mediated cell damage.
Protein-folding stress response. When cells accumulate misfolded proteins — which happens routinely under stress, and pathologically in many neurodegenerative diseases — the ER mounts a coordinated response called the unfolded protein response (UPR). The sigma-1 receptor is involved in regulating this response, helping cells handle misfolding stress more effectively. Loss of sigma-1 function makes cells more vulnerable to protein-misfolding pathologies.
Mitochondrial coupling. Because the sigma-1 receptor sits at the contact zone between ER and mitochondria, it is well-positioned to influence the energy and signalling exchange between them. Sigma-1 activation has been shown to support mitochondrial function under stress, including by helping maintain the integrity of mitochondrial membranes and the efficiency of energy production.
Cellular survival and neuroplasticity. Several lines of evidence suggest that sigma-1 activation promotes the survival of neurons under stress and supports the growth of new synaptic connections. This effect operates on longer timescales than the acute effects of surface-receptor drugs — days and weeks rather than minutes and hours. It overlaps with some of the mechanisms now invoked to explain psychedelic neuroplasticity more broadly.
Modulation of other receptors. As a chaperone-like protein, the sigma-1 receptor influences the activity of various other receptors and ion channels it associates with — including some opioid receptors, some serotonin receptors, and several ion channels. This modulatory role means that activating sigma-1 can change how other neuronal signalling systems behave, even without directly stimulating those systems.
The aggregate picture is of a small protein doing several jobs at once, all of them concerned with how cells cope with stress and adapt to challenge. None of these jobs corresponds to the kind of rapid acute signalling that surface receptors handle. The sigma-1 receptor operates at a different temporal layer of cellular biology — closer to maintenance and resilience than to moment-to-moment activity.
5. Why DMT?
Here is the connection that brings the sigma-1 receptor into the orbit of this site.
In 2009, a paper by Dominique Fontanilla, Arnold Ruoho, and colleagues at the University of Wisconsin reported a striking finding. N,N-dimethyltryptamine — the molecule covered in detail in What is DMT? — bound to the sigma-1 receptor and behaved as an agonist (an activator) at this site. The reported affinity was strong enough to raise the question of whether endogenous DMT could matter at sigma-1 under particular local or stress-related conditions, although whether such concentrations are actually achieved in living human tissues remains unresolved.
The finding mattered for three reasons.
First, it connected DMT to a target that had previously seemed unrelated to the classical psychedelic story. 5-HT2A was, and remains, the principal target for the acute psychedelic experience. Sigma-1 was something else — an intracellular protein with no obvious role in producing visual phenomena or ego dissolution. The Fontanilla paper raised the possibility that DMT might have two pharmacological stories: producing the acute experience through 5-HT2A, and doing something else through sigma-1, on different timescales and with different consequences.
Second, it opened a possible explanation for endogenous DMT. If the trace amounts of DMT detected in human biological fluids — discussed in detail in the Endogenous DMT review — were too small to produce psychedelic effects through 5-HT2A, they might nevertheless be sufficient to engage sigma-1 under the right conditions, where lower concentrations can still matter. This made the puzzle of endogenous DMT slightly more tractable: maybe the body's own DMT does not need to be at psychoactive concentrations to be doing something biologically meaningful, if its target is sigma-1 rather than 5-HT2A.
Third, it connected DMT to the broader sigma-1 research programme. Once DMT was identified as a sigma-1 agonist, the question of what sigma-1 activation does — for cellular stress, for neuronal survival, for protein-folding responses, for mitochondrial function — became a question about what DMT might be doing on longer timescales than the acute experience.
Subsequent work has extended the sigma-1 discussion to DMT-family compounds, including 5-MeO-DMT, especially in cellular and immunological models. The precise importance of sigma-1 binding differs by compound and remains an active research question.

6. The Parkinson's strand
The sigma-1 receptor has acquired particular research interest in one specific clinical context that ARDMT's Field Notes follow closely: Parkinson's disease.
Parkinson's is a neurodegenerative condition in which neurons in specific brain regions — particularly dopamine-producing neurons in the substantia nigra — progressively die. The cellular mechanisms of this death involve, among other things, protein-misfolding stress, mitochondrial dysfunction, and the loss of neuroplastic capacity. All three are processes that sigma-1 activity is known to influence.
Several preclinical studies, beginning in the late 2010s and accelerating through the early 2020s, have demonstrated that sigma-1 agonists can protect dopamine-producing neurons in animal models of Parkinson's disease. The protection appears to operate through multiple mechanisms — stabilising mitochondrial function, supporting the unfolded protein response, promoting neuronal survival under metabolic stress — all of which are aspects of sigma-1's normal cellular role.
This has produced interest in sigma-1 agonists as possible neuroprotective or disease-modifying agents in Parkinson's and related neurodegenerative contexts. Some sigma-1-directed compounds have entered clinical development, although the Parkinson's-specific evidence remains early and uncertain. The compounds under investigation are not DMT itself — pharmacokinetic and regulatory considerations make a controlled psychedelic difficult to develop as a chronic neurological treatment — but synthetic sigma-1 agonists designed to retain the desired cellular effects without producing acute psychedelic experiences.

The relevance for the broader DMT story is significant. If sigma-1 activation is genuinely neuroprotective in Parkinson's, and if DMT is a sigma-1 agonist, then questions about whether DMT itself (or analogues, or related compounds) might have disease-modifying effects in Parkinson's become reasonable rather than fanciful. This is one of the more interesting strands of contemporary psychedelic-related research, partly because it bridges the worlds of psychedelic pharmacology and conventional neurology in a way that few other findings do.
It is worth emphasising that this remains preclinical and early-clinical work. The translation from animal models to human disease modification is notoriously unreliable, and the Parkinson's research landscape is littered with promising preclinical findings that did not survive larger human trials. The sigma-1 strand is interesting; it is not yet a clinical reality.
ARDMT's Field Notes track this strand actively. Search for "sigma-1" or "Parkinson's" in the archive to find specific trial coverage as it appears.
7. The therapeutic horizon
Beyond Parkinson's, sigma-1 agonism has been investigated, with varying degrees of preclinical support, in a range of contexts worth noting briefly.
Depression and mood disorders. Several antidepressant drugs already in clinical use — including fluvoxamine (a common SSRI) — have significant sigma-1 agonist activity in addition to their primary pharmacological actions. There is active debate about how much of these drugs' antidepressant effect is attributable to sigma-1 versus their primary targets. Dedicated sigma-1 agonists are being investigated as potential antidepressants in their own right.
Neuroplasticity and cognitive enhancement. Sigma-1 activation supports neuroplasticity, and several lines of research are exploring whether sigma-1 agonists might be useful in conditions characterised by impaired plasticity — including some forms of cognitive decline, post-stroke recovery, and traumatic brain injury rehabilitation.
Pain and neuropathy. Sigma-1 modulators have been investigated for chronic pain conditions, particularly neuropathic pain that is poorly responsive to conventional analgesics. The mechanisms are not fully understood but appear to involve sigma-1's modulatory effects on other pain-signalling systems.
Anti-cancer effects. Some sigma-1 modulators have shown activity against cancer cell lines, apparently through effects on the cellular stress responses that cancer cells depend on for survival. This is at an early stage and not particularly the subject of clinical work yet.
Neuroprotection more broadly. Beyond Parkinson's, sigma-1 agonists are being investigated for their potential to protect neurons in other contexts — stroke, traumatic brain injury, some forms of dementia. The underlying logic is the same: sigma-1 supports cellular resilience, so activating it may help neurons survive insults that would otherwise kill them.
The aggregate picture, as with harmine in its own primer, is of a target whose biological importance is becoming more apparent as research accumulates, and whose therapeutic potential spans several clinical contexts. Whether any sigma-1-directed therapy reaches widespread medical use in the next decade remains to be seen.
The connection to DMT and the other psychedelic tryptamines through this target is one of the more genuinely interesting strands of contemporary psychedelic pharmacology — not because anyone seriously expects DMT itself to become a Parkinson's medicine, but because the existence of this binding suggests that the body's evolved relationship with DMT-like compounds may be operating at a deeper layer of cellular biology than the acute-experience story alone captures.
8. Is there a legal dimension?
Like the Default Mode Network, the sigma-1 receptor is not a substance and cannot be controlled by drug legislation. It is a protein expressed in human cells, and possession of the protein is, of course, universal among living humans.
The substances that bind to sigma-1 vary widely in their legal status. DMT, 5-MeO-DMT, and several other psychedelic agonists are controlled. Many non-psychedelic sigma-1 ligands — including fluvoxamine, dextromethorphan (the cough-suppressant in many over-the-counter cold remedies), and various research chemicals — are either approved medications or unscheduled. Some are widely available; some are tightly controlled. The legal landscape for sigma-1 pharmacology overall is therefore patchwork, depending on which specific compound is being considered.
What is consistent across jurisdictions is that sigma-1 research itself — investigations of the receptor's biology, its function in disease, its responses to non-controlled compounds — is universally permitted under standard research ethics frameworks. The bottleneck for psychedelic-sigma-1 research is the same bottleneck affecting all psychedelic research: the legal status of the active compounds, not of the target.
9. What people often get wrong
Recurring misconceptions about the sigma-1 receptor and its relationship to psychedelic pharmacology, briefly addressed:
"DMT works by activating sigma-1." Not principally. The acute psychedelic experience produced by DMT operates through 5-HT2A receptors on the surface of cortical neurons. Sigma-1 activation may contribute to longer-timescale effects — neuroplasticity, cellular resilience, possibly some aspects of the experience itself — but the headline psychedelic action is a 5-HT2A story. Treating sigma-1 as the explanation for what DMT does at psychoactive doses overstates what the evidence currently supports.
"Sigma-1 is the receptor for endogenous DMT." This is one hypothesis the field has discussed; it is not established. Endogenous DMT might engage sigma-1; it might engage other targets; it might be doing nothing biologically meaningful at the concentrations the body actually produces. The endogenous DMT question and the sigma-1 question are connected but distinct, and the connection between them remains genuinely open.
"Activating sigma-1 produces psychedelic effects." It does not, at least not in the conventional sense. Many sigma-1 agonists used in research and medicine — including approved drugs like fluvoxamine — do not produce psychedelic experiences. The acute psychedelic state is a 5-HT2A phenomenon; sigma-1 activation does something different.
"Sigma-1 and sigma-2 are related receptors." They are not. Despite the shared "sigma" name, the two were grouped together historically because of overlapping pharmacology in the early days of the field. Molecular characterisation has since established that they are entirely different proteins, encoded by unrelated genes, with different functions and tissue distributions. The naming is a historical accident.
"DMT's binding to sigma-1 proves it's a fundamental endogenous signalling molecule." It proves binding. It does not prove function, role, or evolutionary purpose. The interpretive leap from "DMT binds to this receptor" to "DMT therefore has a fundamental role in human biology" is much larger than the evidence supports and has been a feature of overclaim in popular psychedelic writing for years. The careful version of the claim is that DMT's binding to sigma-1 is consistent with several interesting possibilities about endogenous function; the careful version does not assert that those possibilities are established.
10. What we still don't know
The genuinely open questions in sigma-1 research, briefly:
What the full set of natural endogenous ligands for the sigma-1 receptor is. DMT and related tryptamines are candidates; so are several lipid-based compounds; so are some neurosteroids. Which of these the receptor evolved to respond to, and at what concentrations, remains unresolved.
Whether the body's own DMT actually engages sigma-1 in a functionally meaningful way at the concentrations endogenous DMT actually achieves. This is the question that would, if answered affirmatively, partly vindicate the Strassman-style "DMT has a fundamental endogenous role" hypothesis; it is also the question the field has been unable to settle for fifty years.
Whether sigma-1 agonism in human Parkinson's disease will replicate the neuroprotective effects observed in animal models. The translation from preclinical to clinical results is the standard hard problem of neurology, and the sigma-1 strand is not exempt from it.
How much of the long-term effects of DMT and related psychedelics on mood, anxiety, and cognition is mediated through sigma-1 versus through other mechanisms (5-HT2A-driven neuroplasticity, downstream effects on the default mode network, indirect effects through other receptors). Disentangling these contributions empirically is difficult.
Whether sigma-1 agonism without psychedelic effects can produce some of the therapeutic benefits attributed to psychedelics — that is, whether a non-psychedelic sigma-1 agonist could substitute for psychedelic therapy in some clinical contexts. Several pharmaceutical programmes are betting that the answer is at least partly yes.
What the broader cellular biology of sigma-1 looks like — its complete set of binding partners, its full regulatory network, its role in normal physiology beyond the disease contexts that have driven most of its research attention.
These are live questions. Several of them are likely to have meaningful answers within the next decade. The sigma-1 receptor has gone from pharmacological curiosity to actively studied target in roughly twenty years, and the rate of progress has been accelerating rather than slowing.
11. Closing the Foundations series
This essay closes the ARDMT Foundations series. The six primers together — DMT, Ayahuasca, 5-MeO-DMT, Harmine and the MAOIs, the Default Mode Network, and this one — were designed as a single on-ramp into the rest of the site. A reader who has worked through all six is now in a position they were not in before starting: equipped with the vocabulary, the pharmacological scaffolding, and the conceptual frameworks that the deeper essays and the daily Field Notes assume.
This is the only structured walkthrough on ARDMT. The rest of the site does not hold the reader's hand in the same way — it operates at the level the Foundations primers were preparing you for. With that in mind, a small amount of guidance about where to go next.
If you want to go deeper on a specific molecule. The Field Notes archive at /tag/research/ carries daily coverage of new research as it appears — clinical trial readouts, mechanistic studies, policy developments. Searching the archive for a specific compound name (DMT, ayahuasca, ibogaine, harmine, 5-MeO) will surface the relevant entries. These pieces are written for readers who already have the Foundations vocabulary.
If you want depth rather than breadth. The Pillar Essays — most prominently the Endogenous DMT in Humans review — are the long-form, fully cited deep-dive pieces. They are denser than the primers, longer (often an hour's reading or more), and assume the reader is willing to engage with the actual scientific evidence in detail. The Foundations primers are the orientation; the Pillar Essays are the work the orientation was preparing you for.
If you want to follow the live debates. Several strands of psychedelic research are genuinely contested in 2026 — the endogenous-DMT question, the synthetic-versus-natural-5-MeO-DMT question, the cultural-translation question around ayahuasca, the harmine-as-therapeutic question, the sigma-1-as-mediator-of-long-term-effects question. These are where the field is currently working things out. Field Notes track them as they develop; the Pillar Essays revisit them when enough has changed to warrant a fresh look.
If you want the ethical and legal dimensions. Forthcoming Pillar Essays on UK psychedelic policy and on the cultural-translation questions will treat these areas in detail. In the meantime, the legal sections within each Foundations primer cover the basics.
If you want what isn't here. ARDMT does not cover everything in the psychedelic landscape. Classical psilocybin research, MDMA-assisted therapy, ketamine clinical work, and several other strands are referenced where they intersect with the DMT/sigma-1/MAOI core but are not the site's primary focus. For sustained coverage of those, other resources exist — and the broader Field Notes coverage will sometimes touch on them as relevant.
The Foundations series exists because the rest of the field is hard to read without it. Now that you have it, the rest of the field is open. The work of orienting is done. Whatever you read next is yours to choose.