Exploration
DEEP DIVE #1 Five Hundred Million Years Apart, Touching Through the Glass: The Octopus, MDMA, and the Deep Chemistry of Social Connection

Abstract
In 2018, Eric Edsinger and Gül Dölen published a short paper in Current Biology that posed an apparently absurd question: what happens when you give MDMA to an octopus? The answer — that the famously solitary, sometimes cannibalistic Octopus bimaculoides became markedly more sociable, "hugging" the cages of conspecifics with extensive ventral surface contact — captured public attention as a curiosity. Beneath the headline, however, sits a more substantial scientific claim with implications for evolutionary biology, neuropharmacology, and the philosophy of mind. Edsinger and Dölen argued that the serotonin transporter binding site responsible for MDMA's prosocial effects in humans is essentially identical in the octopus, and that the behavioural circuitry it regulates may therefore have been conserved across more than 500 million years of evolution — including across the radical reorganisation of nervous systems that produced the cephalopod brain on one branch and the mammalian cortex on the other. This article reviews the original study, the critical commentaries it received, and what its findings — and limits — mean for the broader project of understanding consciousness, social experience, and the chemistry that appears to underwrite both.
Introduction
Picture a tank at the Marine Biological Laboratory in Woods Hole. A small California two-spot octopus drifts out of the central compartment of a three-chambered arena and toward the perforated plastic enclosure holding another male — a conspecific it had spent the previous trial studiously avoiding from the opposite end of the tank. It does not retreat. Instead, it drapes its arms across the barrier, presses its mouth-parts and ventral suckers against the mesh, and stays there. In the description Gül Dölen will later give to journalists, it is hugging the cage. Twenty minutes earlier, the animal had been immersed in seawater carrying a low dose of MDMA, absorbed through its gills. More than five hundred million years of evolutionary divergence separate it from the humans watching. Whatever has just happened, both species appear to recognise it as familiar.
Octopuses are, on most counts, very poor candidates for a study of prosocial behaviour. Octopus bimaculoides typically lives alone in a den, treats conspecifics as competitors, predators, or — in the case of males approaching females — potentially lethal threats (Cigliano, 1993; Hanlon and Messenger, 2018). Cannibalism is well-documented across the cephalopods (Ibáñez and Keyl, 2010), and during mating events that do occur, males tend to extend a specialised hectocotylus arm from the maximum distance their morphology allows (Gutnick and Kuba, 2018). Whatever a prosocial octopus might look like, it is not the species' modal phenotype.
This was precisely what made the Edsinger–Dölen experiment interesting. (±)-3,4-Methylenedioxymethamphetamine (MDMA) reliably produces feelings of warmth, connectedness, and reduced threat-perception toward conspecifics in humans (Kamilar-Britt and Bedi, 2015), and similar effects — increased adjacent lying, reduced aggression — have been characterised in rodents (Heifets and Malenka, 2016). The pharmacology is now reasonably well-understood: MDMA binds the serotonin transporter SERT (encoded by SLC6A4), inhibiting reuptake and producing elevated synaptic serotonin alongside downstream effects on oxytocin and dopamine signalling (Liechti, 2014; Heifets and Malenka, 2016). It is worth flagging at the outset that MDMA is properly categorised not as a classical psychedelic but as an entactogen: it acts by elevating synaptic serotonin via reuptake inhibition, whereas the classical psychedelics — psilocybin, LSD, DMT, mescaline — act primarily as direct 5-HT2A receptor agonists. The two classes are pharmacologically distinct, though they share enough overlapping circuitry, and are studied within enough overlapping research programmes, that casual coverage often groups them together.
The question Edsinger and Dölen posed was whether a creature whose last common ancestor with humans was a basal bilaterian living over half a billion years ago — a creature that lacks a cortex, and distributes roughly two-thirds of its half-billion neurons into its eight arms (Hochner, 2012; Godfrey-Smith, 2016) — would respond to this drug as we do.
It did. The interpretive task that follows is harder than it might appear.
1. The Unlikely Subject
If one set out to choose an animal that had evolved intelligence by a different route to vertebrates, one could not do much better than the octopus. The molluscan lineage diverged from the chordate lineage during or just before the Cambrian, with conservative molecular-clock estimates placing the split around 550 to 600 million years ago (Albertin et al., 2015; Schwarz et al., 2024). What evolved on the molluscan branch was, neurologically speaking, a profoundly different solution to the problem of being a complex organism.
Coleoid cephalopods — octopuses, squid, and cuttlefish — possess the largest invertebrate nervous systems known, with Octopus vulgaris carrying approximately 500 million neurons, comparable in raw count to a dog (Hochner, 2012; Mather, 2019). Crucially, only around a third of these neurons reside in the central brain; the remainder are distributed across eight peripheral arm ganglia, each capable of substantial autonomous sensorimotor processing (Hochner, 2012; Godfrey-Smith, 2016). The arms can taste, grip, and explore without direct central oversight; the philosopher Peter Godfrey-Smith (2016) has argued that this distributed architecture demands we rethink the relationship between intelligence, embodiment, and the unity of the self.
The cephalopod central brain itself is organised on principles foreign to vertebrate neuroanatomy. There is no cortex. Instead, the supraesophageal and subesophageal masses are divided into 30 to 40 lobes, with associative regions — most notably the vertical lobe — performing functions analogous to those of mammalian cortex and hippocampus, but arrived at through convergent rather than homologous evolution (Shigeno et al., 2018; Schwarz et al., 2024). The Cambridge Declaration on Consciousness (Low et al., 2012) explicitly named cephalopods alongside mammals and birds as possessing the neurological substrates of conscious experience, an inclusion that remains striking given how fundamentally cephalopod nervous systems differ from our own.
This is the organism Edsinger and Dölen chose. Its evolutionary distance from humans is the entire point: any shared response to a psychoactive compound becomes, by virtue of that distance, a candidate for a deeply preserved mechanism rather than a recent coincidence.
2. The Genomic Foundation
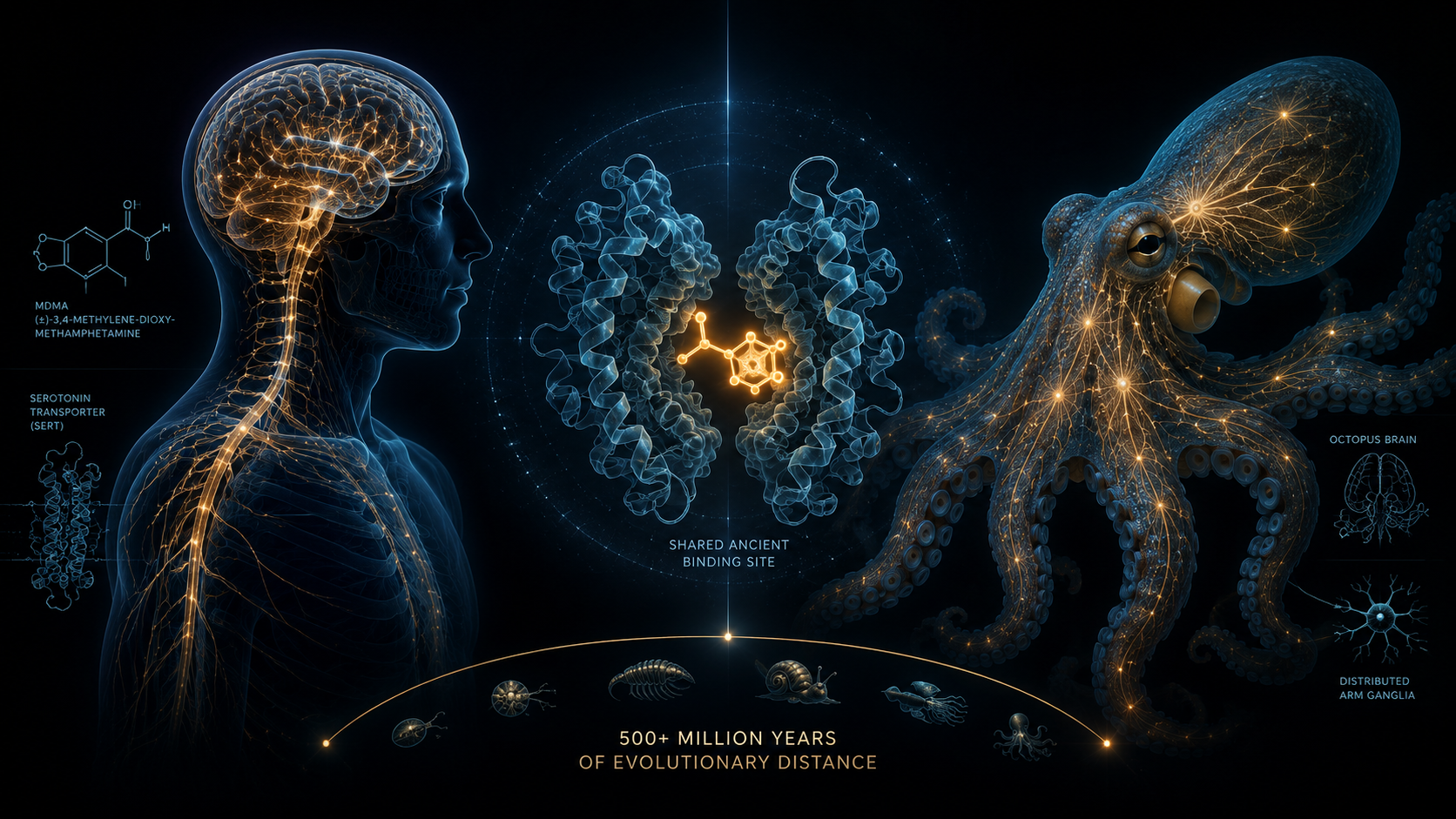
The study's first move was genomic. Building on the 2015 publication of the Octopus bimaculoides genome (Albertin et al., 2015), of which Edsinger had been a co-author, the researchers performed sequence alignments of the octopus orthologues of SLC6A4, the gene encoding SERT, against the human reference (Edsinger and Dölen, 2018).
The binding pocket of SERT is formed by a subset of twelve transmembrane domains, with transmembrane domain six (TM6), and specifically the residues spanning amino acids 333 to 336, providing an overlapping binding site for both endogenous serotonin and exogenous MDMA (Andersen et al., 2011; Edsinger and Dölen, 2018). Residue Ser336 has been implicated in the specific conformational change MDMA produces that distinguishes its effects from those of serotonin alone (Andersen et al., 2011). What Edsinger and Dölen reported was that, in both octopus orthologues of SLC6A4, this functionally critical region showed 100 per cent amino acid identity with the human sequence. More broadly, the binding pocket was substantially more preserved than the full-length protein, suggesting purifying selection operating specifically on the regions that matter for ligand interaction.
This is a strong claim that requires careful framing. Sequence identity does not, by itself, establish functional equivalence: identical residues can sit within nervous systems that route signals to entirely different behavioural outputs. The molecular finding therefore set up, rather than settled, the behavioural question.
3. Methods
Because no behavioural pharmacology protocol existed for octopuses, the researchers had to build one. Rather than injecting MDMA — invasive, stressful, and likely to confound any behavioural readout — they immersed each octopus in seawater containing the compound, allowing absorption through the highly vascularised gill epithelium (Edsinger and Dölen, 2018; Gutnick and Kuba, 2018). The effective dose corresponded approximately to the human low-to-moderate recreational range. Animals were then transferred to clean seawater and tested over a 30-minute window.
The behavioural assay was adapted from the three-chamber social approach test, a paradigm well-established in rodent neuroscience (Yang et al., 2011). A central chamber connected to two flanking chambers: one contained an unfamiliar conspecific under a perforated cage, the other a novel object — initially a Star Wars action figure — under an identical cage. Time spent in each chamber, and the quality of interaction, were scored from video.
Two experiments were run. The first (n = 5) established baseline sex-conditioned preferences in drug-naive animals encountering both male and female caged conspecifics. The second (n = 4) used a within-subjects pre-post design to test the effect of MDMA on approach toward a familiar male.

4. Results
In the baseline condition, O. bimaculoides showed clearly sex-conditioned approach: drug-naive octopuses spent more time in the chamber containing a female conspecific than in the chamber containing a novel object, and conversely tended to avoid the chamber containing a male conspecific, preferring even the object chamber to a male stranger (Edsinger and Dölen, 2018, Figure 3). This pattern is consistent with the species' natural history, in which male–female contact occurs during mating but male–male encounters are typically agonistic, and corroborates earlier behavioural work in the genus (Cigliano, 1993; Hanlon and Messenger, 2018).
The MDMA condition produced a measurable shift. Following exposure, all four animals spent more time in the social chamber with the male conspecific than they had during the pre-trial baseline (Edsinger and Dölen, 2018). More striking, in the authors' qualitative description, was the character of the interaction. Where pre-trial encounters with the caged male had been brief, cautious, and typically conducted at a distance, post-MDMA interactions involved animals approaching the cage directly, draping their arms across its surface, pressing their mouth-parts and ventral suckers against the perforated barrier, and maintaining that contact across extended periods — a posture Dölen has described, in interviews accompanying the publication, as "hugging" the cage (Edsinger and Dölen, 2018, p. 3139; Johns Hopkins Medicine, 2018). The investigators drew an explicit parallel to the increased frequency of affiliative touch reported in human MDMA sessions (Kamilar-Britt and Bedi, 2015).
The combination of the genomic finding — a near-perfectly preserved SERT binding site — and the behavioural finding provided what the authors framed as the first functional evidence that the prosocial effects of serotonergic neurotransmission are conserved across the protostome–deuterostome divide (Edsinger and Dölen, 2018).
5. What the Study Shows — and What It Does Not Show
The paper was published alongside an invited commentary by Gutnick and Kuba (2018), and a more pointed methodological critique appeared the following year from Amodio et al. (2019). Together, these responses mark the boundary between what the data plausibly support and what they do not.
What the data support
The genomic finding is robust. The high conservation of the SERT binding site is a structural fact about two sequenced genomes and is not contingent on the behavioural experiment. There is also a real, measurable behavioural shift: drug-naive octopuses that avoided a caged male conspecific spent more time in that chamber following MDMA exposure, and the direction of the effect matches the predicted serotonergic mechanism. Whatever one thinks of the qualitative gloss, the quantitative shift in approach behaviour did occur.
Where the data run out
Gutnick and Kuba (2018), themselves cephalopod neuroscientists, were broadly enthusiastic but flagged two issues. First, the description of octopuses as uniformly solitary and cannibalistic is itself a simplification of a behaviourally diverse clade: several species, including Octopus tetricus and the larger Pacific striped octopus, exhibit tolerated aggregations and non-aggressive social structures in the wild (Scheel et al., 2017; Rodaniche, 1991; Gutnick and Kuba, 2018). The contrast between "naturally asocial" and "MDMA-induced prosocial" is therefore softer than the headline framing suggests. Second, the immersion delivery method, while elegant, makes precise pharmacokinetic claims difficult; without invasive sampling it is hard to know what plasma concentration of MDMA the octopuses actually achieved.
The more substantive critique came from Amodio, Fiorito, Clayton and Ostojić (2019), writing in Frontiers in Behavioural Neuroscience. They raised three concerns. First, sample size: with four animals in the MDMA experiment, statistical power is necessarily low and individual variability dominates any inference. Second, they identified an issue with the way baseline data had been used: through correspondence with the original authors, they established that for three of the seven animals across the study, data from one trial were used both as the endpoint of Experiment 1 and as the baseline for Experiment 2 — a methodological detail not disclosed in the original publication and one that complicates the assumption of independent observations (Amodio et al., 2019). Third, and most damaging to the qualitative claim, they argued that "extensive ventral surface contact" with a caged stimulus is not a social-specific response in octopuses. The species routinely deploys precisely this exploratory repertoire — arm wrapping, mouth-part contact, sucker engagement — toward inanimate objects during foraging, problem-solving, and play (Fiorito et al., 1990; Kuba et al., 2003; Hanlon and Messenger, 2018). Without an MDMA-condition object chamber demonstrating that this behaviour was preferentially directed at conspecifics rather than at any salient stimulus, the qualitative argument for prosociality is, on their reading, undersupported.
The genomic finding stands. The strong behavioural claim — that this represents the first functional demonstration of evolutionarily conserved prosocial effects — does not yet.
None of this eliminates the possibility that MDMA produces genuinely prosocial effects in O. bimaculoides. What the commentaries do is recalibrate confidence, and point clearly to the next experiment: larger samples, an MDMA-condition object control, and a more granular behavioural ethogram.
6. Why It Resonated, and What It Means
Before unpacking the deeper implications, it is worth pausing on why this paper found such purchase in the public imagination. The viral life of the Edsinger–Dölen study was not merely a function of its octopuses-on-ecstasy headline. The paper sat at the intersection of three currents in contemporary scientific culture, each carrying its own accumulating momentum.
The first was a rapidly expanding popular appreciation of cephalopod intelligence, catalysed by Godfrey-Smith's Other Minds (2016), Sy Montgomery's Soul of an Octopus (2015), and a steady stream of viral natural-history footage demonstrating tool use, escape behaviour, and individual recognition. The second was a wholesale reframing of psychedelic and entactogen pharmacology from drugs-of-abuse to candidate therapeutics, exemplified by the parallel progression of MDMA-assisted psychotherapy through Phase 3 trials for PTSD (Mitchell et al., 2021). The third was a broader philosophical interest — increasingly visible in both academic and trade publishing — in whether the basic infrastructure of social and affective experience is older and more widely shared than vertebrate-centric accounts have allowed. The Edsinger and Dölen paper provided, in a single image, a point of synthesis for all three currents. That cultural salience matters: it explains why a methodologically modest study has been disproportionately influential, and it sharpens, rather than dilutes, the importance of getting the underlying claims right.
6.1 The shared chemistry of social regulation
Serotonin is one of the oldest signalling molecules in the eukaryotic toolkit. It is found across the animal kingdom and even in plants, where it regulates growth and stress responses (Erland et al., 2016). Its role as a neuromodulator predates the divergence of protostomes and deuterostomes; nematodes, flatworms, molluscs, arthropods and chordates all employ serotonergic signalling in roles regulating arousal, feeding, locomotion, and — as Edsinger and Dölen's work suggests — social engagement (Curran and Chalasani, 2012). The transporter machinery MDMA targets is similarly old. The pharmacological space within which psychedelic and entactogen compounds operate is not a recent vertebrate development but a series of perturbations to regulatory chemistry that has been doing roughly similar work since long before brains in the modern sense existed.
This has implications for how we think about psychedelic pharmacology more generally. Classical serotonergic psychedelics — psilocybin, LSD, DMT, mescaline — act primarily at the 5-HT2A receptor, itself a member of a widely preserved family of G-protein-coupled receptors with homologues across the bilaterian lineage (Nichols, 2016). Receptor antiquity does not, of course, guarantee that subjective experiences mediated by it are phenomenologically comparable across species. But it does undercut a certain anthropocentric framing in which psychedelic states are treated as exotic intrusions on an otherwise stable human cognitive baseline. They may instead be perturbations of regulatory systems that have been shaping the affective and social lives of organisms for a very long time.
6.2 The cortex is not a precondition
A persistent assumption in much philosophy of mind, and in some areas of neuroscience, is that the structures necessary for rich conscious experience — and certainly for anything resembling drug-induced affective states — are cortical, or at minimum vertebrate-pallial (Tononi et al., 2016). Cephalopods complicate this assumption considerably. They lack a cortex entirely. Their associative regions, principally the vertical lobe, are organised on different developmental and architectural principles to mammalian pallium, evolved convergently, and yet support a range of behaviours — observational learning, tool use, individual recognition, contingent play, and possibly subjective pain experience — that are difficult to account for without granting some form of integrated experience (Mather, 2019; Schwarz et al., 2024; Crook, 2021).
This argument deserves an immediate counterweight. Behavioural similarity is not subjective similarity. An octopus approaching another octopus under serotonergic modulation does not necessarily imply anything like a human experience of empathy, warmth, or interpersonal bonding. Similar behavioural outputs can emerge from radically different experiential architectures — or, in some philosophical accounts, from no experiential architecture at all (Frankish, 2016). The inference from "this organism's behaviour shifts in the direction ours does, under the same chemical" to "this organism is experiencing what we experience" requires bridging premises that the current data cannot supply. The cephalopod case does not collapse the gap between behaviour and experience; it merely shows that the gap cannot be sealed shut by appeal to neuroanatomy alone.
What the case does establish is more modest but still significant. The structural assumption that affective regulation by serotonergic perturbation requires cortex-like architecture is not safe. The cortex is plausibly sufficient for certain kinds of integrated affective states; it is increasingly difficult to argue that it is necessary. That alone has obvious bearing on how we extend ethical consideration to non-mammalian animals, and on what we should expect from psychedelic studies in invertebrate systems.
6.3 Implications for psychedelic research
The Edsinger and Dölen (2018) study sits within a broader and rapidly expanding programme of psychedelic and entactogen research that has moved, in the past decade, from the margins to the mainstream of neuropsychiatric investigation (Carhart-Harris and Goodwin, 2017; Mitchell et al., 2021). Within this programme, animal models have always played a contested role: critics ask, reasonably, whether rodent head-twitch responses or sociability assays can tell us anything meaningful about the subjective and therapeutic dimensions of human psychedelic experience.
The octopus work shifts this conversation in a small but interesting way. It suggests that the chemical substrate of at least one psychedelic-adjacent compound is sufficiently widely preserved that even an organism on a radically different neural plan responds in a directionally similar fashion. This is, on one hand, an argument for taking comparative pharmacology seriously: a behavioural effect appearing across this kind of evolutionary distance is unlikely to be an artefact of vertebrate-specific quirks. On the other hand, it is an argument for epistemic humility about the relationship between behaviour and experience: directional similarity at the behavioural level admits of multiple interpretations.
For the developing field of psychedelic-assisted therapy, the practical takeaway is modest but not trivial. The pharmacological systems being targeted are not idiosyncratically human; they are old regulatory machinery shared across most of the animal kingdom. Treatments that work through these systems are, in this sense, working with deep biology rather than against it.
Conclusion
Edsinger and Dölen's 2018 study is a small, methodologically imperfect, evocative paper. Its sample sizes are modest, its qualitative behavioural analysis is open to the criticisms Amodio and colleagues subsequently levelled, and its dosimetry is approximate. None of this is unusual for an exploratory study opening a new experimental paradigm in a species for which no behavioural pharmacology infrastructure previously existed.
What survives the methodological scrutiny is the genomic finding and the directional behavioural shift. The serotonin transporter binding site that mediates MDMA's prosocial effects in humans is essentially identical in Octopus bimaculoides, and acute exposure to MDMA produces a measurable change in conspecific approach behaviour in a normally avoidant species. Across the phylogenetic gulf of more than half a billion years, across the divide between organisms with and without a cortex, across the difference between centralised and partially distributed nervous systems, the same chemical lever appears to move something recognisable. What that something is — at the level of subjective experience, at the level of conscious affect, at the level of whatever it is that makes a human on MDMA reach for another person — remains an open question.
It is, however, a question that the octopus has now made it impossible to dismiss as merely a human one.
References
Albertin, C. B., Simakov, O., Mitros, T., Wang, Z. Y., Pungor, J. R., Edsinger-Gonzales, E., Brenner, S., Ragsdale, C. W. and Rokhsar, D. S. (2015) 'The octopus genome and the evolution of cephalopod neural and morphological novelties', Nature, 524(7564), pp. 220–224.
Amodio, P., Fiorito, G., Clayton, N. S. and Ostojić, L. (2019) 'Commentary: A conserved role for serotonergic neurotransmission in mediating social behavior in octopus', Frontiers in Behavioral Neuroscience, 13, p. 185.
Andersen, J., Stuhr-Hansen, N., Zachariassen, L., Toubro, S., Hansen, S. M. R., Eildal, J. N. N., Bond, A. D., Bøgesø, K. P., Bang-Andersen, B., Kristensen, A. S. and Strømgaard, K. (2011) 'Molecular determinants for selective recognition of antidepressants in the human serotonin and norepinephrine transporters', Proceedings of the National Academy of Sciences, 108(29), pp. 12137–12142.
Carhart-Harris, R. L. and Goodwin, G. M. (2017) 'The therapeutic potential of psychedelic drugs: past, present, and future', Neuropsychopharmacology, 42(11), pp. 2105–2113.
Cigliano, J. A. (1993) 'Dominance and den use in Octopus bimaculoides', Animal Behaviour, 46(4), pp. 677–684.
Crook, R. J. (2021) 'Behavioral and neurophysiological evidence suggests affective pain experience in octopus', iScience, 24(3), 102229.
Curran, K. P. and Chalasani, S. H. (2012) 'Serotonin circuits and anxiety: what can invertebrates teach us?', Invertebrate Neuroscience, 12(2), pp. 81–92.
Edsinger, E. and Dölen, G. (2018) 'A conserved role for serotonergic neurotransmission in mediating social behavior in octopus', Current Biology, 28(19), pp. 3136–3142.e4.
Erland, L. A. E., Turi, C. E. and Saxena, P. K. (2016) 'Serotonin: an ancient molecule and an important regulator of plant processes', Biotechnology Advances, 34(8), pp. 1347–1361.
Fiorito, G., von Planta, C. and Scotto, P. (1990) 'Problem solving ability of Octopus vulgaris Lamarck (Mollusca, Cephalopoda)', Behavioral and Neural Biology, 53(2), pp. 217–230.
Frankish, K. (2016) 'Illusionism as a theory of consciousness', Journal of Consciousness Studies, 23(11–12), pp. 11–39.
Godfrey-Smith, P. (2016) Other Minds: The Octopus and the Evolution of Intelligent Life. London: William Collins.
Gutnick, T. and Kuba, M. J. (2018) 'Animal behavior: socializing octopus', Current Biology, 28(19), pp. R1147–R1149.
Hanlon, R. T. and Messenger, J. B. (2018) Cephalopod Behaviour. 2nd edn. Cambridge: Cambridge University Press.
Heifets, B. D. and Malenka, R. C. (2016) 'MDMA as a probe and treatment for social behaviors', Cell, 166(2), pp. 269–272.
Hochner, B. (2012) 'An embodied view of octopus neurobiology', Current Biology, 22(20), pp. R887–R892.
Ibáñez, C. M. and Keyl, F. (2010) 'Cannibalism in cephalopods', Reviews in Fish Biology and Fisheries, 20(1), pp. 123–136.
Johns Hopkins Medicine (2018) Octopuses given mood drug 'ecstasy' reveal genetic link to evolution of social behaviors in humans. Press release, 20 September.
Kamilar-Britt, P. and Bedi, G. (2015) 'The prosocial effects of 3,4-methylenedioxymethamphetamine (MDMA): controlled studies in humans and laboratory animals', Neuroscience and Biobehavioral Reviews, 57, pp. 433–446.
Kuba, M., Meisel, D. V., Byrne, R. A., Griebel, U. and Mather, J. A. (2003) 'Looking at play in Octopus vulgaris', Berliner Paläobiologische Abhandlungen, 3, pp. 163–169.
Liechti, M. E. (2014) 'Effects of MDMA on body temperature in humans', Temperature, 1(3), pp. 192–200.
Low, P., Panksepp, J., Reiss, D., Edelman, D., Van Swinderen, B. and Koch, C. (2012) The Cambridge Declaration on Consciousness. Cambridge: University of Cambridge.
Mather, J. A. (2019) 'What is in an octopus's mind?', Animal Sentience, 4(26), p. 1.
Mitchell, J. M., Bogenschutz, M., Lilienstein, A. et al. (2021) 'MDMA-assisted therapy for severe PTSD: a randomized, double-blind, placebo-controlled phase 3 study', Nature Medicine, 27(6), pp. 1025–1033.
Montgomery, S. (2015) The Soul of an Octopus: A Surprising Exploration into the Wonder of Consciousness. New York: Atria Books.
Nichols, D. E. (2016) 'Psychedelics', Pharmacological Reviews, 68(2), pp. 264–355.
Rodaniche, A. F. (1991) 'Notes on the behavior of the larger Pacific striped octopus, an undescribed species of the genus Octopus', Bulletin of Marine Science, 49(1–2), p. 667.
Scheel, D., Chancellor, S., Hing, M., Lawrence, M., Linquist, S. and Godfrey-Smith, P. (2017) 'A second site occupied by Octopus tetricus at high densities, with notes on their ecology and behavior', Marine and Freshwater Behaviour and Physiology, 50(4), pp. 285–291.
Schwarz, R., Stark, G., Levin, M., Gutnick, T. and Kuba, M. J. (2024) 'Evolution of cephalopod nervous systems', Annual Review of Neuroscience, 47, pp. 79–98.
Shigeno, S., Andrews, P. L. R., Ponte, G. and Fiorito, G. (2018) 'Cephalopod brains: an overview of current knowledge to facilitate comparison with vertebrates', Frontiers in Physiology, 9, p. 952.
Tononi, G., Boly, M., Massimini, M. and Koch, C. (2016) 'Integrated information theory: from consciousness to its physical substrate', Nature Reviews Neuroscience, 17(7), pp. 450–461.
Yang, M., Silverman, J. L. and Crawley, J. N. (2011) 'Automated three-chambered social approach task for mice', Current Protocols in Neuroscience, 56(1), pp. 8.26.1–8.26.16.